文博信息
宋光捷 蔡大伟 | 中国绵羊的起源与扩散研究新进展
摘要: 一、引 言 绵羊(Ovis aries)在动物分类学上属于偶蹄目(Artiodactyla)、洞角科(Bovidae)、羊亚科(Caprinae)、绵羊属(Ovis)。作为人类最早驯化的家养动物之一,绵羊在古代社会发展过程中扮演了一个重要的角色,不仅为古代人类社会提供了许多稳定的、高品质动物蛋白来源和皮、毛、奶等产品,还在宗教祭祀和占卜等方面影响着民族文化的形成与发展。 ...
一、引 言
绵羊(Ovis aries)在动物分类学上属于偶蹄目(Artiodactyla)、洞角科(Bovidae)、羊亚科(Caprinae)、绵羊属(Ovis)[1]。作为人类最早驯化的家养动物之一,绵羊在古代社会发展过程中扮演了一个重要的角色,不仅为古代人类社会提供了许多稳定的、高品质动物蛋白来源和皮、毛、奶等产品,还在宗教祭祀和占卜等方面影响着民族文化的形成与发展。绵羊最早的驯化证据可以追溯至大约距今11000年前,其驯化的起点位于新月沃地[2]。考古证据表明,在公元前9000~前8000年,从土耳其中部到伊朗西北部的地区,当地人类已经开始进行绵羊管理[3];例如,安纳托利亚中部和东南部的遗址中出土绵羊的圈养迹象和对年轻雄性宰杀行为[4]。公元前7500年之后,对年轻雄性绵羊的宰杀和与驯化相关的形态变化(例如体型变小)在新月沃地(包括安纳托利亚中部)广泛出现,因此伊朗西北部到安纳托利亚可能是绵羊驯化的中心[5]。
绵羊有7个野生近缘种,分别亚洲摩弗伦羊(Asiatic Mouflon,Ovis gmelinii)、欧洲摩弗伦羊(European Mouflon,Ovis orientalis musimon)、盘羊(Agarli,Ovis ammon)、加拿大盘羊(Bighorn,Ovis canadensis)、阿尔卡尔羊(Urial,Ovis vignei)、雪羊(Snow sheep,Ovis nivicola)和大白羊(Thinhorn,Ovis dalli)[6]。而绵羊起源于何种野羊一直存在争议。其中亚洲摩弗伦羊、盘羊和阿尔卡尔羊被认为与家绵羊血缘关系最近或者对绵羊驯化有贡献[7],因此关于绵羊起源的问题十分复杂,而亚洲摩弗伦羊被认为最有可能是家养绵羊的野生祖先[8]。切萨(Chessa)等研究表明欧亚大陆和非洲的绵羊经历了两次独立驯化事件:第一次是人类为了获取肉食,驯化了亚洲摩弗伦羊等野生羊;第二次是古代人类为了获取羊毛资源,驯化并筛选培育高产毛量的绵羊[9]。
驯化后的绵羊通过人群迁徙、贸易等方式被传播到邻近地区,包括欧洲、北非和中亚,随后绵羊扩散到世界各地[10]。绵羊向欧洲传播主要是大约10000年前,绵羊到达了安纳托利亚中部,再由地中海和多瑙河谷两条主要的传播路线到达欧洲[11];此外,还有另一条通往欧洲的传播路线:通过俄罗斯高加索地区到达北欧[12]。绵羊可能通过两条路线进入北非:第一条是穿越地中海盆地到达非洲;第二条是穿越西奈半岛,然后向下到达红海并穿越红海[13]。美洲和澳大利亚的绵羊主要是近代由欧洲和非洲引入[14]。根据分子生物学研究,绵羊向亚洲的传播主要有三条路径:①蒙古高原是绵羊扩散的第二个中心,驯化中心的绵羊沿高加索和中亚地区迁徙到蒙古高原,并从此处到达中国北方、西南和南亚次大陆;②驯化中心的绵羊从中东通过阿拉伯地区传入南亚次大陆;③通过古代海上贸易路线,绵羊从近东引入南亚次大陆[15]。
在中国,许多新石器时代早期考古遗址中出土了大量的羊的遗骸和陶羊、羊圈等与绵羊驯化有关的遗存。在河南新郑裴李岗遗址、陕西半坡遗址、陕西元君庙灰坑中都曾出土羊的骨骼。但由于未对其进行形态测量和种属鉴定,所以不知道是绵羊还是山羊,也不知道是驯化的还是野生的[16]。袁靖先生认为中国最早出现的家养绵羊是在距今5600~5000年前的甘肃省和青海省一带,然后向东由关中地区传播进入中原地区,到距今4500~4000年前黄河中下游地区出土家养绵羊遗骸比较丰富,确证龙山文化时期养羊业已经有了一定的发展[17]。胡松梅认为我国遗址中确定为家养绵羊的时代,最早在西阴文化时期(距今6000~5200年)的泉护村遗址[18];其传播路径为:绵羊到达蒙古高原后分为两支,一支分沿黑河向南到达河西走廊一带,另一支向东到达中国榆林地区,再由北向南传播到中原地区[19]。综上所述,中国古代家绵羊应该是由国外引进的,但具体传入时间和传入路径尚未明确。本文拟通过结合动物考古和分子考古学研究成果进行综述,使人们对中国古代家绵羊起源、驯化和扩散的过程有综合、全面的认识。
二、中国绵羊的动物考古学研究
动物考古学是研究绵羊起源与扩散的有利方法之一,对此,袁靖先生提出了一系列方法用于鉴定绵羊和研究绵羊的起源问题,这些方法包括形体测量、病理现象、年龄结构、性别特征、数量比例、考古现象、食性分析等[20]。中国尚有很多遗址出土大量的羊骨,但由于骨骼形态特征不明显、骨骼质量较差、骨骼结构相似等原因,导致动物考古学者无法分辨出土的羊骨是绵羊还是山羊,因此常以“羊”或“绵羊/山羊”来代替[21]。一些野生牛科动物在骨骼形态等方面也与绵羊有相似之处,这也会使得研究者无法对其进行准确的鉴定,导致我们不能清楚了解绵羊起源和扩散的情况[22]。此外,出土羊骨件数过少也会导致研究者无法判断绵羊是否为家养。例如,杨苗苗等对河南渑池笃忠遗址(仰韶中晚期)出土的一件下颌骨进行鉴定,确定其为绵羊,并根据牙齿磨损级别判断死亡年龄为6~8岁,但由于仅出土一件下颌骨,不能确定其是否为家养[23]。
目前已经有部分遗址出土的羊骨经过系统骨骼形态学鉴定确认为家养绵羊,如杨春对西岔遗址(商周时期)出土的绵羊骨骼进行了系统鉴定与分析,鉴定为家养绵羊,标本数共有346件,最小个体数为64[24];李谅对长宁遗址(齐家文化)出土羊骨进行分析,根据使用羊骨祭祀、随葬和占卜鉴定为家养绵羊,同时也根据最小个体数判断出土绵羊和山羊的比值[25]。此外,动物考古学可以通过判断绵羊死亡年龄和骨骼部位占比来研究家绵羊的利用方式。如博凯铃对陶寺遗址出土绵羊牙齿萌出和磨蚀情况发现超过40%的绵羊被宰杀时年龄超过4~6岁,说明当时开发绵羊的主要目的是羊毛等副产品而不是肉食[26]。李倩对河南新砦遗址各期出土绵羊骨骼部位发现率进行分析研究,发现率较高的部位是下颌骨,其次为肩胛骨和髋骨,说明下颌骨密度较大且肉量较少,先民较少利用,家犬也较少啃食,因此保存较好;而肩胛骨和髋骨这两个部位肉量较多,表明先民存在食肉的利用方式[27]。
目前出土的羊遗骸中,除已确定的绵羊外,尚有许多无法确定其为绵羊或山羊,为了解决这一问题,还需要依赖DNA分析进行分子考古学研究来获取更丰富的证据。
三、中国绵羊的分子考古学研究
1.绵羊的线粒体DNA研究
线粒体DNA(mtDNA)因为遵循母系遗传模式、具有多拷贝数并且具有比核基因更高的突变率,使得其可以很好地用于解决系统发育关系和群体多样性[28]。线粒体DNA也因此被广泛应用到绵羊起源扩散的研究中。
线粒体DNA单倍型研究应在逻辑上反映绵羊驯化的地理进程。在绵羊mtDNA的特定区域中,人们最常关注的是控制区(D-Loop)区和细胞色素b编码区域[29]。目前可以确定五个绵羊存在母系世系(A、B、C、D和E)(图一)。A世系和B世系是最早发现也是最主要的两个绵羊母系世系,其中世系A大多数分布亚洲,世系B中大部分分布在欧洲[30]。2005年,郭等对分布在中国6个地方的绵羊品种进行了mtDNA分析,发现了一个比A、B两个世系更具遗传变异性的新世系C[31]。佩德罗萨(Pedrosa)等在研究近东地区的绵羊时也发现了世系C,表明家绵羊的起源比之前认为的更复杂,至少发生了三次独立的绵羊驯化事件[32];C世系绵羊主要分布在葡萄牙、高加索、中东和亚洲地区。2006年塔皮奥(Tapio)等通过分析北高加索地区的绵羊控制区序列发现了一个母系世系(世系D)[33]。2007年梅多斯(Meadows)等通过分析土耳其八个品种的绵羊的线粒体控制区和细胞色素b的序列,发现了新的世系E[34]。世系D和世系E较为稀有,迄今为止仅在高加索和土耳其的绵羊中发现。2013年德米尔奇(Demirci)等人在对土耳其家绵羊线粒体进行系统发育分析时,发现了一个绵羊新的世系(世系X)[35];但是到目前为止,世系X仅有这一项报道,没有后续研究证实它的存在[36]。综上所述,线粒体DNA序列分析已经确定家养绵羊的母系起源有5个,绵羊的5个世系在不同地区的频率分布如下。
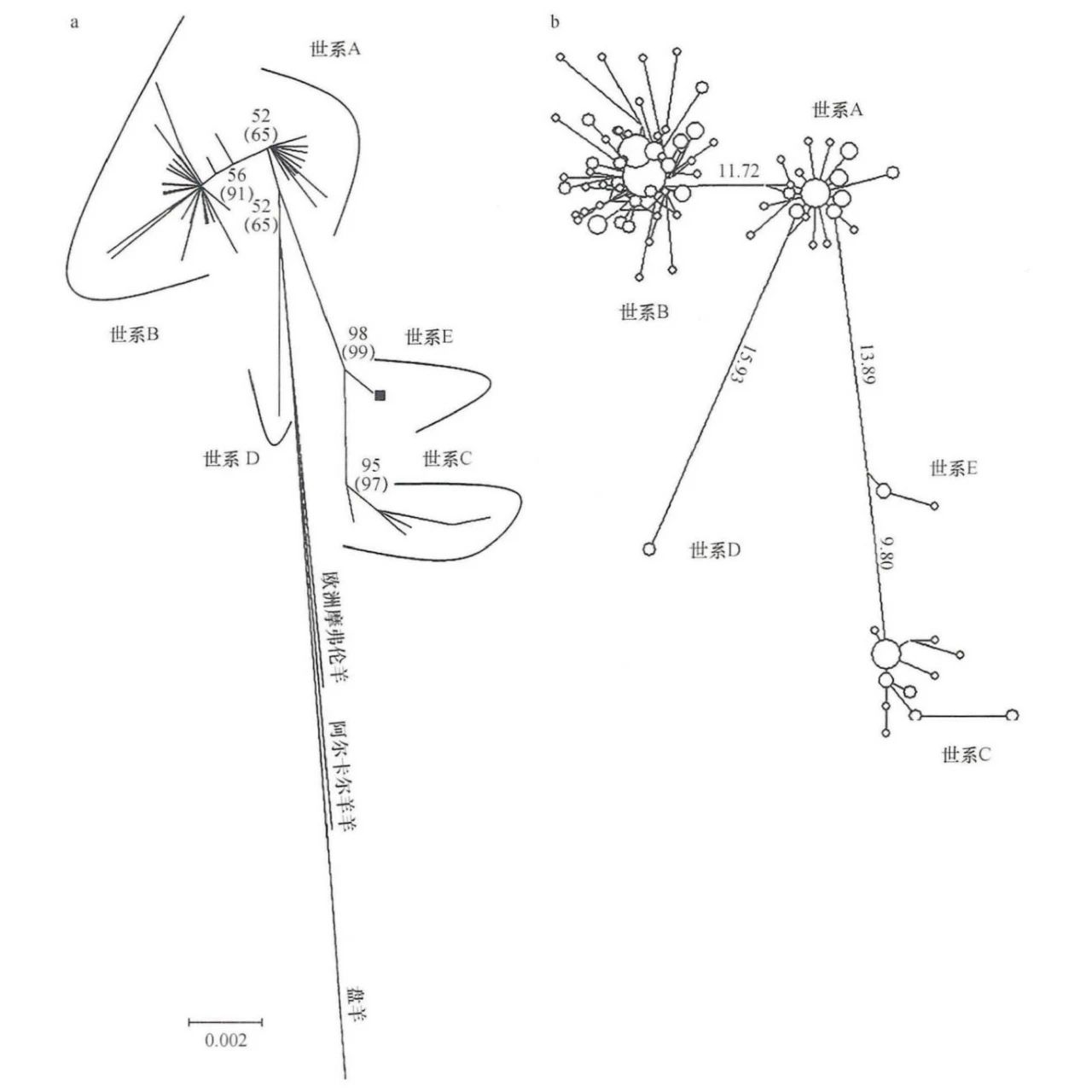
图一 绵羊母系世系分类示意图[34]
a.邻接树 b.中介网络图
中国绵羊的起源与扩散的时间、地点、路线以及中国是否为绵羊的一个驯化中心一直以来是研究人员关注的重点。2006年,陈善元等对中国13个地区的19个绵羊品种进行了线粒体DNA控制区分析,发现世系A存在两个亚分支,推测绵羊A系发生了两次独立的驯化事件,暗示中国也是绵羊的驯化中心之一,说明绵羊的驯化地点不仅限于近东地区,还发生在其他地区[37]。2009年,韩璐等对内蒙古春秋战国时期24个古代绵羊进行控制区和细胞色素b基因片段进行分析,揭示了中国绵羊有3个不同的母系起源,还发现中国古代绵羊与中国现代绵羊的遗传结构极为相似,这暗示了2500年前中国绵羊的遗传结构就已经趋于稳定,并对现代绵羊的基因有很大的贡献[38]。2010年,蔡大伟等通过分析研究中国7个遗址出土的45个古代绵羊线粒体控制区序列,发现在距今4000年前青铜时代早期,中国古代绵羊只有A和B两个世系,其中A世系占统治性地位;在距今2500年前的春秋战国时期,绵羊群体中出现了世系C;从距今2500年前开始,占比最低的D、E两个世系开始出现;此外,使用古代绵羊序列与从基因数据库(Genbank)中选取现代绵羊和野生绵羊序列构建的系统发育网络图表明古代绵羊和现代绵羊之间存在遗传联系,进一步支持世系A起源于东亚,而野生绵羊中摩弗伦羊聚在B世系中,说明摩佛伦羊与家绵羊的关系较近;其他野生绵羊如源羊、盘羊和大角羊等与中国绵羊没有直接的血缘关系[39]。2013年,赵二虎等分析16个中国绵羊品种线粒体DNA D-loop区序列,发现中国除西南山区仅有A、B世系外,其他地区均有A、B、C三个世系,未发现D、E世系[40]。2017年,俄广鑫等分析中国西藏156个绵羊的线粒体DNA D-loop区序列,暗示阿尔卡尔羊和盘羊对西藏绵羊的起源有贡献,西藏绵羊中未发现D、E世系,说明西藏绵羊不是起源于中东地区[41]。
绵羊的五个世系是根据控制区片段和细胞色素b基因定义的,但是这两个片段占整个基因组的比例不到12%,突变是在线粒体全基因组的各个组成部分之间以不同速率积累所得,基于线粒体DNA短区域生成的系统发育树,其分支拓扑缺乏统计稳健性;其次,在追溯家绵羊的野生祖先时,野生绵羊和家绵羊可能会杂交产生后代,因此使用线粒体全基因组可以揭示更多信息[42]。随着测序技术的发展,线粒体全基因组分析对绵羊母系起源的研究起到至关重要的作用。2011年,梅多斯等通过使用10个家养绵羊和三种野生绵羊(欧洲摩弗伦羊、阿尔卡尔羊和盘羊)的线粒体全基因组序列数据构建系统发育树发现:盘羊和阿尔卡尔羊均不是家养绵羊的母系祖先,与之前的研究结果不同的是,与阿尔卡尔羊相比,盘羊与家绵羊的关系更为密切;欧洲摩弗伦羊与世系B聚在一簇,说明欧洲摩弗伦羊不是传统意义上的野生羊,可能是经过驯化后再次返野[43]。2015年,布拉希(Brahi)等对西非家绵羊全基因组测序研究,揭示西非绵羊来自欧亚大陆,而最初西非驯化的绵羊品种彼此之间已经进行了基因杂交,并与欧洲品种混合[44]。同年,桑娜(Sanna)等对家养绵羊和野生绵羊线粒体全基因组进行系统发育分析,揭示阿尔卡尔羊和盘羊分别在距今89万和111万年前与家绵羊分化;绵羊的五个世系分为两个主要分支(A、B、D和C、E)并在30万年前发生分化,其中世系D在24万年前最先被分化,A和B世系在17万年前分化,C和E在12万年前分化[45]。同年,吕锋骅等分析整个欧亚大陆家养和野生绵羊的线粒体全基因组,通过对比与结合线粒体DNA片段分析结果,揭示出:①五个家绵羊世系中共同的母系祖先生活在大约距今79万年前的中更新世时期,比它们的驯化时间早得多;②绵羊的祖先在末次盛冰期(26000~19500年前)之前经历了种群的快速扩张;③绵羊在6800~4500年前在史前人类活动的影响下沿不同路线经历了两次迁徙(第一次:A和B世系;第二次:C世系),构建了横跨欧亚大陆的现代绵羊原始母系遗传结构,而中国华北地区可能是C世系绵羊的一个起源地;④蒙古高原是绵羊扩散的第二中心,来自驯化中心的绵羊经过高加索和中亚地区并穿过该地区到达中国北部、西南地区(A、B和C世系)和印度次大陆(B和C世系);⑤世系A从中东通过阿拉伯传入印度次大陆[46]。
2.绵羊的Y染色体研究
线粒体DNA信息仅通过母系遗传,因此对线粒体DNA的分析结果不能完全代表整个绵羊种群的历史动态。而Y染色体由于只能由雄性亲本遗传给雄性子代,因此Y染色体的分析可以作为研究雄性世系遗传与进化的一个有效的分子标记[47]。其中Y染色体的雄性特异区域(MSY)因其不与X染色体重组、突变率比mtDNA低等特点可用于绵羊起源、驯化以及遗传多样性的研究[48]。目前利用Y染色体进行绵羊父系起源的研究较少,均为利用Y染色体单倍型进行父系起源的研究。使用Y染色体单倍型研究绵羊父系起源和种群扩散的两个关键分别是在MSY上的性别决定基因(SRY)中的8个SNP位点(oY1-oY8)标记和微卫星SRYM18标记[49]。
2006年,梅多斯等通过分析世界范围内65个品种的458只家绵羊,发现A-oY1的频率最高(71.4%);对微卫星SRYM18序列分析发现大角羊、瘦角羊与欧洲摩弗伦羊、家绵羊有明显不同;结合基因型数据确定了11个绵羊Y染色体单倍型,它们至少代表两个独立的世系,其中单倍型H6在全球分布最广泛,其他单倍型在世界范围内严格遵守地理分布规律;该研究得出现代绵羊存在欧洲驯化中心和中东驯化中心两个父系起源[50]。2009年,梅多斯等通过微卫星标记SRYM18,首次发现17个Y染色体单倍型H1~H17;此外,分析结果显示摩弗伦羊是唯一与家羊共享单倍型的野生物种[51]。2013年,费伦查科维奇(Ferencakovic)等分析东亚得里亚海的9个家养绵羊品种的159只公羊和21只亚洲摩弗伦羊的Y染色体单倍型,首次发现了单倍型H18,这提供了土耳其少数公羊(单倍型H12)与一只埃塞俄比亚公羊(单倍型H9)之间的联系[52]。2014年,张敏等对来自欧亚大陆北部59个品种的663只公羊的两个SNP和一个Y染色体微卫星的变异进行了分析,发现了新的单倍型H19并发现该地区存在7个单倍型(H4、H5、H6、H7、H8、H12和H19);欧亚大陆北部以H6为主;而之前报道在欧洲品种中不存的H4,在一个欧洲品种中被检测到;之前确定为欧洲品种所独有的H7,在两个中国品种、一个布里亚特品种和两个俄罗斯品种中被检测到;此外,仅在土耳其品种中检测到的H12在中国品种中也有发现;在中国西北部和东部,H6是占主导地位的单倍型,同时也有部分单倍型H4和H8存在[53]。2015年,王玉涛等对中国11个品种的绵羊Y染色体进行分析,得出中国绵羊群体存在7个单倍型(H4、H5、H6、H7、H8、H9和H12),并且在中国家绵羊中首次发现了2个新单倍型(Ha和Hb);其中Ha在1个品种中出现,Hb在3个品种中出现[54]。
2020年,邓娟等对179个公羊个体进行分析,首次识别绵羊Y染色体特异区493个多态位点,确定了四个世系(y-HA、y-HB、y-HC和y-HD)(图二)。通过对芬兰和爱沙尼亚绵羊古DNA分析,发现北欧古代绵羊群体和现代绵羊群体的遗传连续性。此外该研究揭示了家养绵羊的多次独立扩张,这可能与不同产品的各种表型性状和育种目标相关[55]。
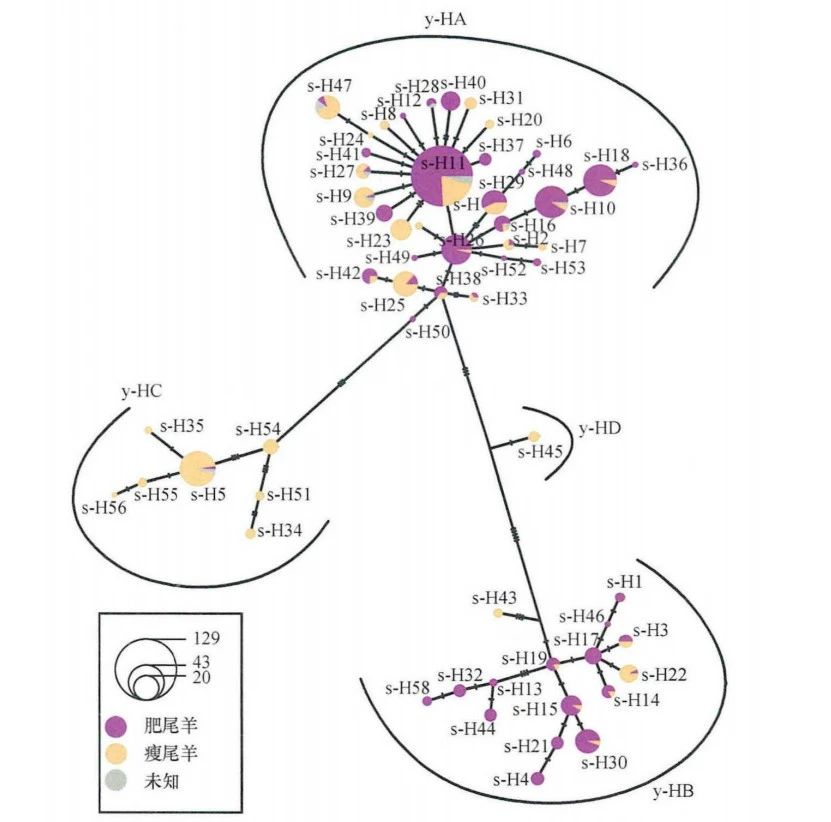
图二 基于绵羊Y染色体特异区的中介网络图[56]
虽然绵羊Y染色体的研究已经取得了一定的进展,但此前绵羊缺乏Y染色体参考序列,这妨碍了Y染色体分子标记在研究绵羊起源与扩散应用。2021年,李冉等构建了第一个绵羊Y染色体序列(共10.6Mb),并通过775个Y-SNP将世界范围的家绵羊分为了三大世系:HY1a、HY1b和HY2;其中HY1b地理分布最广、频率最高,HY1a仅在欧洲种群中发现,HY2主要在中国、伊朗和非洲种群中发现;此外,该研究还发现HY2与HY1a和HY1b的遗传关系较远,而与伊朗摩弗伦野羊聚为一支,暗示HY2代表一个独立的绵羊世系且来源于伊朗摩弗伦野羊的渗入[57]。
3.绵羊的核基因组研究
近年来,随着二代测序技术的飞速发展,绵羊的分子生物学研究进入到了核基因组时代,相比于线粒体DNA和Y染色体研究,核基因组可以更全面、精准地从时间和空间上把握古代家绵羊起源和驯化的问题研究。2014年由以中国科学院昆明动物所和深圳华大基因研究院领衔的国际研究小组历时五年的努力,公布了参考基因组序列,为绵羊全基因组研究奠定了基础[58]。
2017年,赵永欣等通过1000多个家养绵羊和野生绵羊的全基因组数据研究了中国羊的基因组多样性;群体基因组分析结合考古记录和历史民族人口统计数据揭示了中国绵羊起源、二次扩张的遗传特征以及东亚游牧的居住模式和早期畜牧的扩张。该研究显示近东地区的绵羊经高加索和中亚地区,在5700~5500年前到达蒙古高原,在5000~3000年前,随着羌人的扩张,中国绵羊开始在黄河上游和中游地区传播,并沿着狄羌人从北到西南的迁徙路线在2600~2000年前到达了青藏和滇黔高原;该研究还推测了两波肥尾羊进入中国北方的迁移潮,这在很大程度上与12~13世纪穆斯林回族祖先向东迁移和蒙古人南迁相一致。此外,还发现了盘羊向家绵羊基因渗入的迹象,绵羊群体之间广泛的历史混合、对尾巴类型和其他性状的强烈人工选择,反映了中国古代游牧社会的各种繁殖策略[59]。
2018年,胡小菊等对西藏986个绵羊的全基因组序列、线粒体DNA和Y染色体进行研究,并检测到参与缺氧和紫外线信号通路(HIF-1通路、HBB和MITF基因)以及关于角的大小和形状的形态特征相关基因(RXFP2);该研究发现盘羊向西藏绵羊基因组中有关于氧运输系统和形态表型基因相关的基因渗入;基因组分析与考古遗址出土证据相结合得出:西藏绵羊是距今约3100年前伴随着人群迁徙从中国北方扩散到青藏高原东北部,随后在距今1300年前向西南扩张到达青藏高原中心[60]。
2021年,泰勒等将核基因组、线粒体DNA分析与动物考古学、ZooMS和牙骨质分析等方法相结合,分析了吉尔吉斯斯坦南部奥比什(Obishir)V遗址出土的绵羊遗骸,发现了早在8000多年前费尔加纳河谷山区已经开始使用绵羊,说明绵羊到达中亚内陆的时间可能远早于之前研究得出的时间[61]。
同年,尤尔特曼(Yurtman)等分析了从旧石器时代到新石器时代晚期的古代安纳托利亚中部和西部绵羊的核基因组和线粒体全基因组数据,并与现代绵羊、中亚新石器时代以及青铜时代绵羊比较;通过分析古代核基因组数据,发现与亚洲品种相比,安那托利亚新石器时代绵羊在基因上与现代欧洲绵羊最接近,线粒体世系也支持这一结论。相比之下,青铜时代绵羊对现代亚洲绵羊表现出更高的遗传亲和力。以上结果表明:在今天的绵羊群体中观察到的东西遗传结构在公元前6000年就已经出现了,这暗示了在西南亚有多个绵羊的驯化事件或早期的野生渗入。此外,我们发现古代安纳托利亚绵羊在基因上不同于所有现代绵羊,表明欧洲和安纳托利亚家绵羊的基因库自新石器时代以来发生了重塑[62]。
同年,陈泽慧等分析了家养绵羊及其野生近缘物种的全基因组数据,发现北美野生绵羊(如雪羊和大白羊等)对阿尔卡尔羊、亚洲摩弗伦羊和欧洲摩弗伦羊有基因渗入;其中欧洲摩弗伦羊可能是由欧洲已灭绝的羊和野化后的家养绵羊杂交产生的。此外,该研究揭示了绵羊及其祖先亚洲摩弗伦羊基因组中与驯化相关的基因之间的差异,最终导致了家绵羊和野生祖先之间的形态、生理和行为的差异(如PAPPA、NR6A1、SH3GL3、RFX3和CAMK4等与形态、免疫能力、繁殖及生产性状有关的基因和NEURL1、PRUNE2、USH2A和PAG11、PAG3等与神经反应、神经发生、听觉能力和胎盘生存能力相关的基因)。这加深了我们对亚洲和欧洲摩弗伦羊的起源以及家绵羊驯化特征的理解[63]。
四、结 语
家养动物的起源、驯化与扩散是人类文明产生和发展的重要环节,也是人类活动与自然环境相互作用的重要事件。本文通过梳理和总结关于家养绵羊起源和扩散问题的研究成果,旨在对中国家养绵羊的研究进展有清晰的认识,为后续的分析与研究提供更多的科学依据。
根据中国绵羊遗骸出土的地点和时间,动物考古学家推测中国家养绵羊可能是在仰韶文化晚期出现在中国的甘青地区和陕北地区,在龙山文化早期进入中原地区;而DNA研究则显示绵羊在距今5700~5500年前经自蒙古高原进入中国,并扩散到黄河中上游地区。这表明中国绵羊起源和扩散的路线还存在较大的争议。近年来,随着分子生物技术和全基因组测序水平的发展,关于绵羊的线粒体全基因组、Y染色体和核基因组研究取得了更多的突破;目前绵羊基因组研究已经应用到与绵羊形态、生产性状、免疫能力等相关的基因研究中,研究人员已经成功筛选出与绵羊毛型、体型、繁殖以及与气候适应相关的基因。这些基因与绵羊的驯化息息相关,通过与绵羊野生近缘物种的比较以及基因渗入的研究,可以反映出人类对绵羊驯化过程中的严格筛选,了解古人在驯化过程中的选择策略。目前在关于研究中国绵羊起源与扩散较多的是分析现代家绵羊的全基因组数据,而对古代样本全基因组数据研究相对较少,后续研究人员可以更多地关注中国古代绵羊的全基因组数据,这样可以更全面、精准地获得有关的信息,进一步解决关于绵羊起源和扩散的问题。
*本文注释从略,阅读全文请参照原刊。
作者:宋光捷(吉林大学考古学院);蔡大伟(吉林大学考古学院,吉林大学边疆考古研究中心,吉林大学-西蒙菲莎大学生物考古联合实验室)
原文刊于:《边疆考古研究》第35辑
责编:昭晣 韩翰
